芒の役割とその消失
 野生イネの多くは種子の先端に「芒(のげ)」とよばれる突起状の構造物を持ちます(図1)。これは種子を包む頴(モミと同義)の中央の維管束が伸長してできた器官であり、長いものでは数十センチにもなります。
野生イネの多くは種子の先端に「芒(のげ)」とよばれる突起状の構造物を持ちます(図1)。これは種子を包む頴(モミと同義)の中央の維管束が伸長してできた器官であり、長いものでは数十センチにもなります。
芒の表面には鋸歯状の細かい棘が形成されており、鳥獣による食害から種子を保護する役割や、人の衣服や動物の毛にからまって種子の拡散を補助する役割があると言われています。しかし、農業を行う上では収穫時の妨げになるとして栽培化の過程で選抜、除去された形質と考えられています。
本研究室ではこの芒という形質に着目し、その遺伝子を単離・解析することにより、イネの栽培化においてどのような分子進化が起きているのかを明らかにしようとしています。
芒の形成に関わる遺伝子
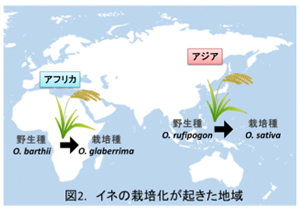
 イネは異なる2ヶ所で独立して栽培化され、アジアでは野生種Oryza rufipogon からO. sativaが、アフリカでは野生種O. barthiiからO. glaberrimaが誕生しました(図2)。本研究室では、これまでに無芒のコシヒカリ(O. sativa)染色体背景に有芒のO. rufipogonの染色体断片が置換した染色体断片置換系統(CSSLs: Chromosome Segment Substitution Lines)を作出し(ref. 1)、その中でコシヒカリの第4染色体の一部がO. rufipogonの染色体に置換した系統と、第8染色体の一部が置換した系統において芒が形成されることを見いだしました。この結果から、O. rufipogonの第4、第8染色体に芒形成遺伝子(それぞれAWN4とAWN8と命名)が存在することが明らかとなりました。AWN4については2013年に中国のグループによって、An-1遺伝子であるということが同定されました(Luo et al., 2013)。
イネは異なる2ヶ所で独立して栽培化され、アジアでは野生種Oryza rufipogon からO. sativaが、アフリカでは野生種O. barthiiからO. glaberrimaが誕生しました(図2)。本研究室では、これまでに無芒のコシヒカリ(O. sativa)染色体背景に有芒のO. rufipogonの染色体断片が置換した染色体断片置換系統(CSSLs: Chromosome Segment Substitution Lines)を作出し(ref. 1)、その中でコシヒカリの第4染色体の一部がO. rufipogonの染色体に置換した系統と、第8染色体の一部が置換した系統において芒が形成されることを見いだしました。この結果から、O. rufipogonの第4、第8染色体に芒形成遺伝子(それぞれAWN4とAWN8と命名)が存在することが明らかとなりました。AWN4については2013年に中国のグループによって、An-1遺伝子であるということが同定されました(Luo et al., 2013)。
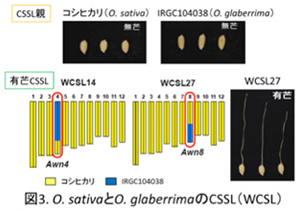
一方、興味深いことに、コシヒカリに無芒の栽培種 O. glaberrimaを交雑したCSSLsにおいても、第4、第8染色体の断片が置換した系統(WCSL14、WCSL27)において芒が確認されました(図3)。この結果はO. glaberrimaは芒形成に対して機能型のAWN4、AWN8を保持しているにも関わらず芒を消失した、すなわちAWN4及びAWN8とは別の要因、別の遺伝子によって芒を消失したことを示唆しています。
これらの結果から、イネは異なる土地で栽培化され、ともに芒を喪失したものの、その過程においてはそれぞれ異なる遺伝子の機能変化が選抜されていると推測されます(ref. 2)。
栽培化された形質において同じ表現型でありながら、異なる遺伝子が選抜されたという例はこれまでに報告が無く、新規の知見として注目しています。今後は芒形成に関わる遺伝子を同定し、芒の栽培化を分子レベルで解明するとともに、栽培化過程における遺伝子制御ネットワークを明らかにしていく予定です。
Reference 1
“Development and evaluation of chromosome segment substitution lines (CSSLs) carrying chromosome segments derived from Oryza rufipogon in the genetic background of Oryza sativa L.” Breed Sci. 63(5):468-75.(2014)
Reference 2
“Independently occurred mutations in independent genes cause awnless phenotype in Oryza sativa and Oryza glaberrima.” in preparation